Constituant tout ou partie du système caulinaire de certains cormophytes, ces organes, le plus souvent souterrains et gorgés de réserves, contribuent à la conservation des espèces. D'une résistance particulière, ils supportent la mauvaise saison en vie ralentie, alors que les parties aériennes ont pu disparaître totalement. C'est sous forme de rhizomes, de tubercules ou de bulbes, que les
géophytes invisibles sous nos latitudes pendant la période hivernale, n'en restent pas moins en vie. En se ramifiant, puis en se fragmentant, ces tiges particulières assurent aussi une multiplication végétative active.
Les rhizomes
Les rhizomes sont des tiges plus ou moins plagiotropes dans la majorité des cas, plus rarement orthotropes, portant des feuilles écailleuses et des racines adventives hypotones. Les rhizomes produisent à la belle saison des rameaux orthotropes aériens feuillés et florifères, ou seulement des feuilles également aériennes, comme les frondes des fougères par exemple. Suivant le nombre et la taille des entre noeuds, on distinguera des rhizomes longs de plusieurs mètres, minces et à croissance rapide, des rhizomes moyennement longs, d'un diamètre supérieur, à croissance moins rapide, et des rhizomes plus ou moins hypertrophiés, à entre noeuds courts et à croissance relativement lente. Variable d'une espèce à l'autre, la profondeur à laquelle croissent les rhizomes est relativement constante pour une espèce donnée. Cette propriété explique en partie la répartition des espèces dunaires rhizomateuses du littoral en fonction de l'importance de l'ensablement local.
 Fig. 16 : rhizomes : A rhizome cespiteux chez Sarracenia sp., B rhizome traçant chez Cynodon dactylon, C rhizome de fougère (Polypodium sp.) portant directement des feuilles et non des tiges feuillées comme dans les deux cas précédents
Fig. 16 : rhizomes : A rhizome cespiteux chez Sarracenia sp., B rhizome traçant chez Cynodon dactylon, C rhizome de fougère (Polypodium sp.) portant directement des feuilles et non des tiges feuillées comme dans les deux cas précédents
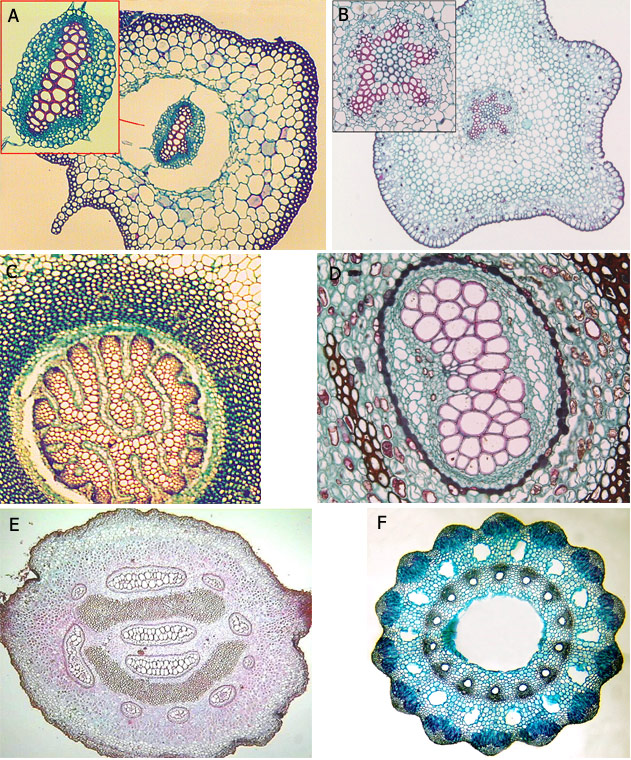
Les tuberculesLes tubercules sont des renflements localisés de la tige principale, ou de ses ramifications. Ils sont généralement souterrains et peuvent avoir plusieurs origines : il existe des tubercules de rhizomes (
Alocasia), des tubercules de stolons (
Solanum tuberosum) et des tubercules hypocotylaires (
Raphanus sativus).
 Fig. 17 : tubercules : A tubercule rhizomateux (Colocasia), B tubercules de stolons (Solanum tuberosum), C tubercules hydrocotylaires (Raphanus sativus)
Fig. 17 : tubercules : A tubercule rhizomateux (Colocasia), B tubercules de stolons (Solanum tuberosum), C tubercules hydrocotylaires (Raphanus sativus)
 Fig. 18 : trois étapes de la tubérisation de l'hypocotyle de radis (c cotylédons, f feuille, h hypocotyle, z.p. zone périphérique de l'hypocotyle déchirée par la tubérisation de la région interne)
Fig. 18 : trois étapes de la tubérisation de l'hypocotyle de radis (c cotylédons, f feuille, h hypocotyle, z.p. zone périphérique de l'hypocotyle déchirée par la tubérisation de la région interne)
Les bulbesLes bulbes sont des tiges le plus souvent orthotrope, larges et plus courtes que les rhizomes, et portent des feuilles en général bien développées, et dont les parties souterraines sont non chlorophylliennes. Suivant l'importance relative de la tige et des feuilles, on distinguera des bulbes caulinaires et des bulbes foliaires. Les
bulbes caulinaires sont constitués d'une tige hypertrophiée enveloppée seulement par des bases foliaires minces et membraneuses. Ils sont denses, et qualifiés de solides. Les
bulbes foliaires ont les réserves situées dans les feuilles sessiles renflées attachées sur le plateau qui est une tige souterraine, réduite et contractée. Les bourgeons axillaires produisent parfois des bulbilles, comme chez l'ail.
 Fig. 19 : bulbes : A bulbe caulinaire (Allium cepa), B bulbe foliaire (Allium sativum)
Fig. 19 : bulbes : A bulbe caulinaire (Allium cepa), B bulbe foliaire (Allium sativum)
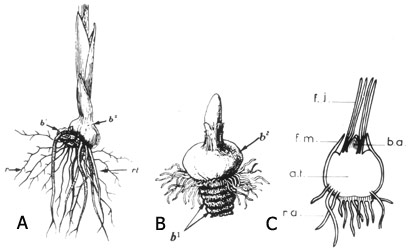 Fig. 20 : bulbes d'Iridaceae : A bulbe rhizomateux plagiotrope de Gladioulus (b1 bulbe de l'an dernier, b² bulbe de l'année), B et C bulbe solide orthotrope de Crocus vernus et de Crocus sativus en coupe longitudinale (r racines adventives ordinaires, r.t. racines adventives tractrices, f.j. jeunes feuilles, f.m., bases membraneuses d'anciennes feuilles, b.a. bourgeon apical, a.t. axe tubérisé, r.a. racines adventives)
Fig. 20 : bulbes d'Iridaceae : A bulbe rhizomateux plagiotrope de Gladioulus (b1 bulbe de l'an dernier, b² bulbe de l'année), B et C bulbe solide orthotrope de Crocus vernus et de Crocus sativus en coupe longitudinale (r racines adventives ordinaires, r.t. racines adventives tractrices, f.j. jeunes feuilles, f.m., bases membraneuses d'anciennes feuilles, b.a. bourgeon apical, a.t. axe tubérisé, r.a. racines adventives)
Les cormes
Un corme est une partie souterraine gonflée de la base d'une tige dressée. Il porte des feuilles et de bourgeons axillaires squamiformes. À la fin de la saison de croissance, les parties aériennes meurent. Avec le retour de conditions favorables, un bourgeon axillaire, rarement plusieurs, se développe près de l'apex en une nouvelle pousse en utilisant le matériel de réserve alimentaire dans le bulbe de l'an passé. La nouvelle année verra apparaitre un nouveau corme à sa base.
 Fig. 21 :corme chez Amorphophallus sp.
Fig. 21 :corme chez Amorphophallus sp.