La fin de la phase d'accroissement cellulaire donne des cellules fortement vacuolisées possédant des plastes structurés et une paroi primaire relativement fine. Les cellules parenchymateuses ne dépassent pas ce stade morphologique de différenciation, leur spécialisation se trouvant au niveau des vacuoles et des plastes. Mais d'autres cellules continuent leur processus de différenciation, et deux possibilités se présentent.
Le cas le plus simple est celui ou toutes les cellules suivent la même voie de spécialisation : elles formeront un
tissus homogène.
Le second cas est celui où mes cellules suivent des voies de spécialisation différentes : le tissus en formation sera
hétérogène. Ainsi, les épidermes renferment des cellules de revêtement, des cellules stomatiques, des cellules tectrices, certaines protectrices, d'autres sécrétrices. Le phloème contient des cellules criblées conductrices, des cellules compagnes, des cellules parenchymateuses auxquelles s'ajoutent souvent des fibres de soutien, des cellules à cristaux, parfois des laticifères, etc.
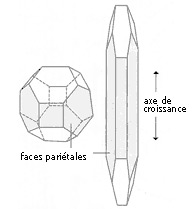
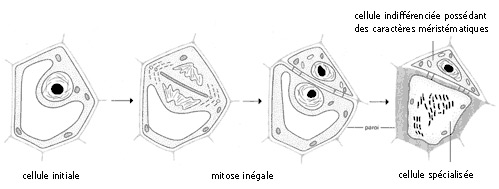 Fig. 5 - Conséquence d'une mitose inégale chez une cellule en voie de différenciation.
Fig. 5 - Conséquence d'une mitose inégale chez une cellule en voie de différenciation.
Si dans les tissus animaux, la spécialisation est souvent acquise par l'élaboration et l'accumulation de protéines spécifiques, chez les végétaux ce phénomène ne s'observe que rarement (cas des cellules criblées du phloème), et la spécialisation est souvent acquise par l'élaboration de parois cellulaires très caractéristiques. C'est ce que l'on nomme
différenciation pariétale.
De primaire, la paroi deviendra secondaire par ajout de nouvelles strates cellulosiques denses et de composés particuliers. Ces composés donnent des propriétés nouvelles aux cellules qui les synthétisent. L'élaboration de lignines, polyphénols incrustant certaines parois, est l'exemple type de cytodifférenciation végétale.
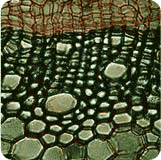
Le cas de la lignification Les tissus ligneux, apparus avec les Cormophytes, permettent le renforcement important des tissus de soutien des végétaux et est responsable de leur port érigé, arbres et arbustes en particulier. La lignine ne se produit normalement que dans le xylème et le sclérenchyme, mais, à la suite d'événements particuliers environnementaux (blessures, attaques parasitaires ou mycosiques...), d'autres tissus peuvent aussi en produire. Inversement, des tissus producteurs de lignine peuvent en ralentir fortement l'élaboration en cas de conditions défavorables : les organes en manque de lumière sont étiolés et ne produisent presque pas de lignine. L'ensemble des cellules semble donc aptes à produire de la lignine, mais, en conditions normales, seules quelques-unes une le font effectivement. L'observation de la lignification dans des conditions environnementales contrôlées a montré que : la lumière, par action sur certaines enzymes, est simulatrice, tout comme la sécheresse, alors que l'humidité est inhibitrice.
Un autre exemple de différenciation pariétale est rencontré chez les cellules destinées à devenir des cellules épidermiques. Situées en contact avec le milieu extérieur aérien, ces cellules se caractérisent par la présence d'une cuticule, couche interface entre le végétal et le milieu extérieur, lui assurant une protection contre les agressions externes et une évaporation d'eau non contrôlée. Il a été montré que la cellule épidermique en cours de spécialisation acquiert l'ensemble des enzymes nécessaires à l'élaboration de la cuticule.