La combinaison des caractères primitifs de la fleur et ceux, très évolués, des organes végétatifs fait des Cactaceae une famille particulièrement intéressante pour les botanistes. Les phytosociologues s'intéressent à sa survie dans des conditions d'extrême sécheresse, et les spécialistes de l'évolution étudient les ressemblances avec des xérophytes appartenant à d'autres familles. Pour les taxonomistes, cette famille pose problème car elle semble encore évoluer et ne peut figurer parmi les spécimens secs d'herbiers. Enfin, sous la pression des collectionneurs et des revendeurs, de nombreux genres ou espèces ont été crées de façon parfois assez artificielle.
On dénombre quatre sous-familles, dont la définition illustre bien le fait rare que la famille fait perdurer de nos jours la complète représentation de son histoire phylogénétique. L'hybridation naturelle est assez fréquente, et est une vraie force pour l'évolution des cactées : de nombreux taxas semblent en effet avoir leur origine dans l'hybridation d'espèces parentes polyploïdes.
La classification utilisée ici repose en grande partie sur les travaux de F. Buxbaum.
La sous-famille des
Pereskioideae (
Pereskia...) regroupe des arbres, des arbustes ou des lianes subsucculents aux feuilles caduques saisonnièrement mais bien developpées, aux tiges droites dépourvues de glochidies, et aux graines noires dépourvues d'arille. Hors floraison, leur relation avec le reste de la famille n'est pas vraiment apparente.
La petite sous-famille des
Maihuenioideae est proche de la précédente, et est limitée à l'Argentine et au Chili.
La sous-famille des
Opuntioideae se caractérise par des arbres, des arbustes ou des sous-arbrisseaux aux feuilles rapidement caduques, présentes sur les jeunes pousses et les fleurs, de forme coniques ou cylindriques, aux tiges plus ou moins cylindriques et pourvues de glochidies, et aux graines couvertes d?un arille pâle et dur.
La sous-famille des
Cactoideae groupe de nombreux genres aux feuilles absentes ou vestigiales et rapidement caduques, aux tiges très succulentes, sphériques à cylindriques ou colonnaires, dépourvues de glochidies, aux graines noires ou brunes non enveloppés par un arille dur. Cette sous-famille, regroupant la majorité des genres, est subdivisée en neuf tribus.
Les Leptocereae ont un port colonnaire souvent ramifié, aux tiges minces munies d'aréoles équipées d'aiguillons ou de duvet (Leptocereus, Armatocereus...).
Les Browningieae ont un port arbustif ramifié, et des rameaux végétatifs très épineux pourvus d'aréoles laineuses (Browningia...).
Les Pachycereae ont un port arborescent, colonnaire, ramifié ou non, avec des tiges devenant côtelées portant des aréoles épineuses ou laineuses.
Les Hylocereae regroupent des genres au port varié, parfois épiphyte, aux tiges rondes et souvent articulées portant des aréoles épineuses, pubescentes ou glabres et aux fleurs parfois zygomorphes (Eriocereus, Epiphyllum, Selenicereus, Hylocereus, Rhipsalis, Schlumbergera...).
Les Cereae (Cereus, Carnegiea, Echinocereus, Echinopsis...) sont des plantes à prédominance colonnaire, avec généralement des tiges articulées aux côtes peu nombreuses, ou des plantes de petite taille fleurissant à partir d'anciennes aréoles.
Les Trichocereae sont représentées par des arbustes parfois ramifiés, dressés ou prostrés, colonnaire ou globuleux, aux fleurs quelquefois zygomorphes (Trichocereus, Cleistocactus, Echinopsis, Rebutia, Leocereus...).
Les Notocacteae sont des sous-arbrisseaux globuleux ou aplatis, rarement colonnaires (Notocactus...).
Les Cacteae (Echinocactus, Ferocactus, Coryphantha, Mammilaria...) sont principalement des plantes de petite taille avec des tiges non articulées aux cotes nombreuses, fleurissant à partir des jeunes aréoles.
Les Cactaceae, parfois érigées en un ordre indépendant (Cactales ou Opuntiales), sont considérées par Wettstein, Emberger, Mangenot, Cronquist, comme faisant partie intégrante des
Caryophyllales (Centrospermales) et placées à proximité des
Aizoaceae et
Didiereaceae. Cette opinion s'appuie sur les similitudes des caractères morphologiques, embryologiques, palynologiques, mais aussi sur la présence de pigments de type bétalaïnes et non pas d'anthocyanes. L'existence de plastes particuliers dans les vaisseaux libériens confirme cette inclusion parmi les Caryophyllales. Ces plastes offrent un cristalloïde central et un anneau périphérique de filaments protéiniques. Il est encore admis que les Cactaceae dérivent des
Phytolaccaceae ou d'un groupe très proche de cette famille. À l'intérieur de la famille, les
Pereskia, aux tiges bien développées, aux feuilles succulentes, aux fleurs terminales parfois groupées en inflorescences et ne possédant pas de glochidies, sont considérées comme les plus primitives, alors que les formes globuleuses (
Mamillaria) sont estimées les plus évoluées.


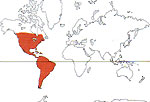 Les cactus sont principalement des plantes des régions chaudes et semi-desertiques des deux continents américains. Seul le genre epiphyte Rhipsalis a colonisé certaines régions d'Afrique, de Madagascar et du Sri Lanka sans intervention humaine, car ses graines sont dispersées par les oiseaux. D'autres genres, comme Opuntia, se sont naturalisés depuis longtemps dans diverses régions tempérées chaudes à tropicales d'Afrique, d'Eurasie et d'Océanie, mais ils ont été cette fois introduits par l'homme, et, dans certaines régions, sont devenus de véritables pestes végétales. La prolifération des Opuntia a été parfois si exceptionnelle qu'il a fallu lutter contre leur envahissement en Australie par l'introduction de prédateurs spécifiques, en l'occurrence un papillon argentin (Cactoblastis cactorum) dont les chenilles minent les raquettes.
Les cactus sont principalement des plantes des régions chaudes et semi-desertiques des deux continents américains. Seul le genre epiphyte Rhipsalis a colonisé certaines régions d'Afrique, de Madagascar et du Sri Lanka sans intervention humaine, car ses graines sont dispersées par les oiseaux. D'autres genres, comme Opuntia, se sont naturalisés depuis longtemps dans diverses régions tempérées chaudes à tropicales d'Afrique, d'Eurasie et d'Océanie, mais ils ont été cette fois introduits par l'homme, et, dans certaines régions, sont devenus de véritables pestes végétales. La prolifération des Opuntia a été parfois si exceptionnelle qu'il a fallu lutter contre leur envahissement en Australie par l'introduction de prédateurs spécifiques, en l'occurrence un papillon argentin (Cactoblastis cactorum) dont les chenilles minent les raquettes.















































































































































